



2010 kavli prize in Neuroscience
2010 Kavli
Prize in
Neuroscience

The Norwegian Academy of Science and Letters has decided to award the 2010 Kavli Prize in Neuroscience to Richard H. Scheller, Thomas C. Südhof and James E. Rothman.



"For discovering the molecular basis of neurotransmitter release."
Committee Members
- Jon Storm-Mathisen (Chair), University of Oslo, Norway
- Tobias Bonhoeffer, Max Planck Institute of Neurobiology, Germany
- Jean-Pierre Changeux, Institut Pasteur, France
- Eric Kandel, Columbia University, USA
- Charles F. Stevens, Salk Institute, USA
Citation from the Committee
The question of how nerve cells communicate with one another has been a central problem in modern neuroscience since the pioneering studies of Cajal, Dale, and Sherrington. By the 1980s, it was well established that nerve cells communicate through the process of chemical synaptic transmission at specialized contacts called synapses. Electron microscopic studies revealed that the presynaptic terminal of the neuron transmitting the information is filled with synaptic vesicles, small organelles containing thousands of molecules of a chemical neurotransmitter. During an action potential, calcium influx into the presynaptic terminal triggers the fusion of synaptic vesicles with the plasma membrane, leading to the release of transmitter through the process of exocytosis. Over the past thirty years, Richard Scheller, Thomas Südhof, and James Rothman have used a creative multidisciplinary set of approaches to elucidate the key molecular events of neurotransmitter release. Moreover, their work demonstrates that neurotransmitter release represents a special case of the fundamental cell biological process of membrane trafficking.
Richard Scheller has used a combination of biochemistry, molecular biology, and cell biology to identify several key synaptic vesicle and plasma membrane proteins involved in fusion of the neurotransmitter-containing vesicles with the membrane of the presynaptic terminal. In particular, he characterized the first synaptic vesicle membrane associated protein, v-SNARE or VAMP, and the first plasma membrane associated target proteins, t-SNAREs or syntaxin and SNAP- 25. Using physiological assays, Scheller demonstrated the importance of these proteins for exocytosis.
Thomas Südhof used powerful biochemical and molecular biological approaches to identify other important synaptic vesicle proteins. He discovered that one of the vesicle membrane proteins, synaptotagmin, had separate calcium and phospholipid binding domains, suggesting it had a key role in transmitter release. Südhof then made use of the emerging power of mouse genetics to delineate the functional role of a number of these vesicle proteins including the role of synaptotagmin, which he demonstrated to be the critical calcium sensor for rapid neurotransmitter release.
James Rothman developed a cell-free assay system to analyze the basic cell-biological processes that mediate membrane trafficking. He identified two soluble proteins (NSF and SNAP) that are important for vesicular transport and membrane fusion in non-neural cells. Remarkably he found that these proteins, when exposed to brain extracts, formed a complex with the vesicle protein VAMP and two plasma membrane proteins, syntaxin and SNAP-25, precisely the proteins that Scheller had identified earlier. The tertiary complex of one vesicle protein, or v-SNARE, with two target membrane proteins, or t-SNAREs, is fundamental not only to transmitter vesicle fusion but to all forms of membrane fusion.
Together these revolutionary insights have given us a fundamental understanding of the molecular basis of neurotransmitter release.
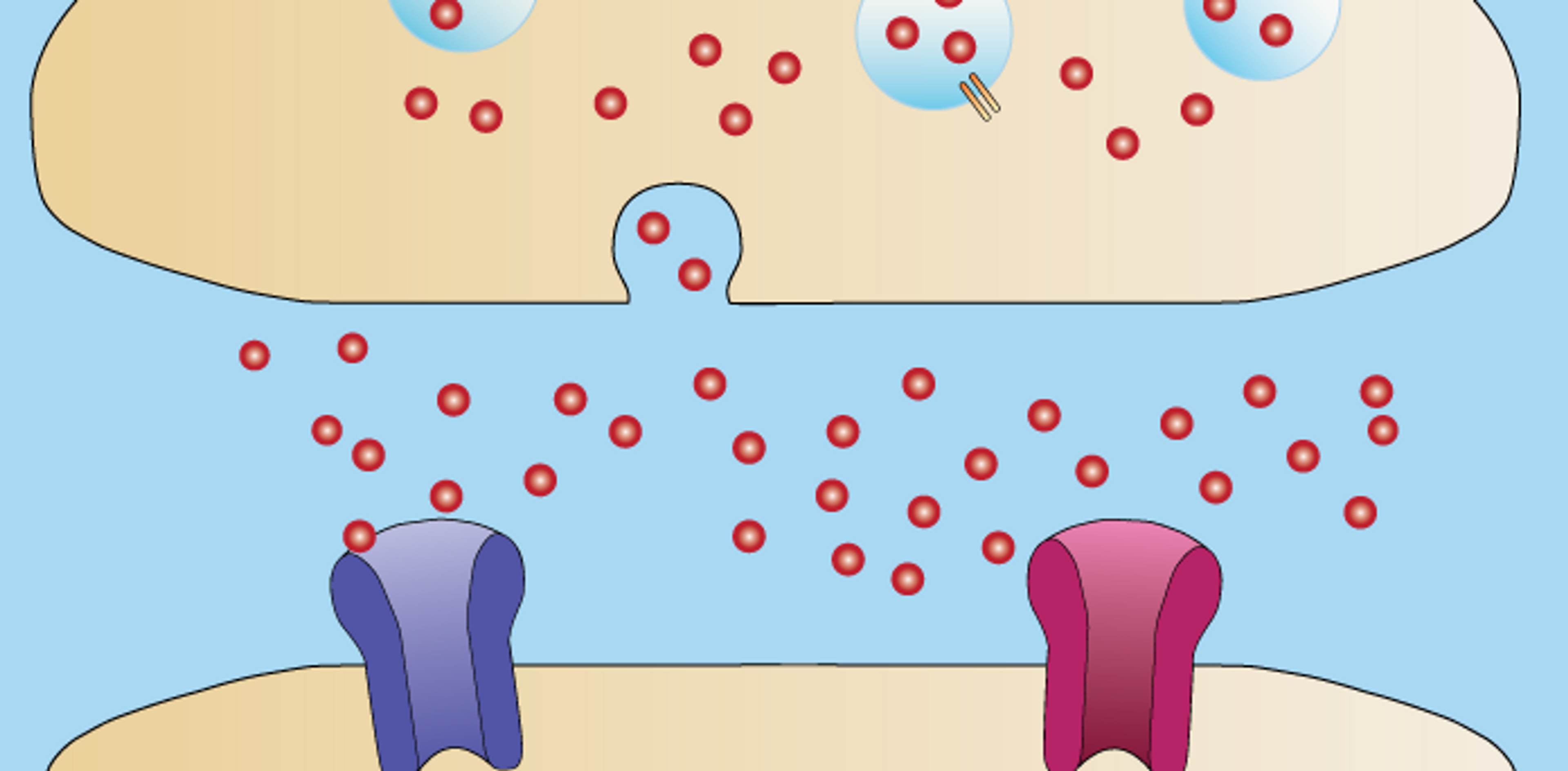
Synapse-Neurotransmitter (Photo credit: Rcchang16, CC BY-SA 4.0)
The 2010 Neuroscience Kavli Prize explained
The 2010 Kavli Prize for Neuroscience has gone to Richard Scheller, Thomas Südhof, and James Rothman who have taken understanding of the nervous system to a new level. Working independently in different laboratories across the U.S., Scheller, Südhof, and Rothman have revealed in exquisite detail the molecules that govern the transfer of signals from one nerve cell to another at special connection points called synapses.
The molecules assemble into a machine that provides an on/off switch for nerve signalling. Without such controls, we would not be able to function – how we think, feel, and respond to everything around us depends on the precise control of signalling across synapses. Over the past 20 years, Scheller and Südhof have focused on the nervous system, identifying key molecules that control the release of chemicals called neurotransmitters at nerve synapses – the tiny junctions where the long extensions of one nerve cell, or neuron, form connections with another. The neurotransmitters act as batons in a relay, transferring signals between neurons in the brain and throughout the body.
Rothman came from a different background in cell biology to discover first serendipitously, and then with extensive follow-up work, that the same molecules identified by Scheller and Südhof also regulate the transfer of material between compartments in cells, as well as secretion by the process of exocytosis, thus showing the much broader significance of these proteins to biology in general.
“The reason for awarding the prize is that they have, in a fascinating and ingenious way, disentangled what happens molecularly at the synapse. It’s one of the first examples in neuroscience where we really understand what single molecules do, how they interact, and what happens if one of them is not there. All of those molecular details are understood to enormous precision in this process which is completely fundamental to the nervous system,” says Prize Committee member Tobias Bonhoeffer of the Max Planck Institute of Neurobiology in Munich, Germany.
“There are many diseases that are diseases of the synapse: schizophrenia, depression. Most psychiatric disorders involve synaptic transmission one way or another. So to be able to intervene and combat these diseases you need to understand the molecular underpinnings of this. And these three people took the study of communication between synapses and brought it from a physiological to a molecular level,” says committee member Eric Kandel of Columbia University, New York.
Until the prizewinners began their work, very little was known about how neurotransmitters were released at synapses. Neuroscientists knew from physiology experiments on whole neurons that thousands of molecules of neurotransmitter are released from nerve endings when a nerve impulse, or "action potential," arrives. These form a chemical relay system, changing the likelihood that another "receiving" neuron sends a new action potential along its fibers (after integrating all the inputs it gets).
At the same time, there is a low level of spontaneous activity at synapses whereby neurotransmitter is released occasionally, producing a kind of background "noise" in terms of neuronal signals. The molecules identified by Scheller, Südhof, and Rothman keep a tight rein on this "noise" to ensure that the system does not become overwhelmed, or that rapid neuronal firing does not occur at the wrong time.
With light microscopy, and then electron microscopy, investigators could define the structure of synapses: where nerve endings come into close proximity with the surface of another neuron, the membrane of the "sending" neuron on one side of the synapse – known as the pre-synaptic membrane – is separated by a tiny gap, the synaptic cleft, from the "post-synaptic" membrane belonging to the "receiving" neuron. By diffusing across this cleft, neurotransmitters convey the signal from one membrane to the other.
Electron microscopy had also given researchers a glimpse of the tiny bubble-like containers called synaptic vesicles that hold thousands of molecules of neuro-transmitter close to the pre-synaptic membrane. There usually are around 10 or so vesicles attached to the pre-synaptic membrane at any one moment. When an action potential arrives one vesicle, on average, forms an opening through which neurotransmitter molecules escape out into the synaptic cleft. With new nerve impulses arriving at intervals of just 10 or 100 milliseconds in some circumstances, other vesicles quickly follow, discharging their contents and refilling, to ensure a steady supply of neurotransmitter.
But while microscopy and electrical recording could provide a broad picture of the structure and timing of neurotransmitter release, no one knew how vesicles reached the pre-synaptic membrane, or the nature of the invisible "threads" that pulled the two together, causing them to fuse and form the opening, or fusion pore. And, critically, what prevented all the assembled vesicles from disgorging their contents together at the wrong time?
In the early 1980s, researchers had used biochemistry to extract vesicles from neurons and analyze their contents, including monoclonal antibodies to identify proteins that were part of the vesicle structure. But the real molecular advance came in the late 1980s when, first Scheller while at Stanford University, then Südhof at the University of Texas Southwestern at Dallas, started to clone and sequence the genes encoding these proteins and to investigate their role in vesicle fusion with the pre-synaptic membrane and neurotransmitter release.
For source material, Scheller first obtained vesicles from the electric organ of the marine ray Torpedo, then switched to rat brain vesicles – the same material used by Südhof. The two teams (Südhof also partly in collaboration with Reinhard Jahn) purified, cloned and defined the functions of virtually all the key proteins involved in neurotransmitter release. Among those first defined by Scheller was the first of what were later called the v-SNARE proteins (the "v" denoting their association with vesicles), and the first of a group of proteins in the pre-synaptic membrane later called the t-SNARE, which the v-SNARE proteins dock into (the "t" standing for "target").
Scheller called the v-SNARE protein that he first cloned in 1988 from Torpedo VAMP-1. Südhof then cloned the rat homologue a year later and called it synaptobrevin. In 1992, Scheller isolated the first t-SNARE and called it syntaxin, based on the Greek “putting together in order."
Scheller also provided the first evidence that a calcium sensing protein called synaptotagmin – discovered earlier by Südhof – was the switch for neurotransmitter release, and proposed in a paper in Science in 1992 that the v-SNARE and t-SNARE proteins combine with synaptotagmin into a three-dimensional scaffold on which other molecules assemble, to form the machinery that initiates vesicle fusion and neurotransmitter release. Synaptotagmin detects the sudden and brief rise in calcium concentration that occurs when an action potential arrives and allows fusion to proceed – all in a fraction of a millisecond.
“This arrangement would ensure that exocytosis occurs both at restricted sites and with an extremely rapid time course,” wrote Scheller (Science, vol 257, 10 July 1992, p258).
In 1993, Rothman dramatically confirmed and extended this idea. He was by this time at the Sloan-Kettering Institute, New York, and had worked on different cell types to see how they released proteins into their surroundings by exocytosis. This involved the movement through the cell interior of vesicles similar in appearance to those of neurons and their fusion with the plasma membrane – which Rothman described as being like “the coalescence of two soap bubbles” – and emptying of their contents. Rothman had demonstrated that the transport and fusion of vesicles involved specific recognition between vesicle-associated molecules, including proteins called NSF and SNAPs (soluble NSF attachment proteins). Others including Randy Schekman had discovered NSF to be important for vesicle trafficking and secretion in yeast, leading Rothman to argue that these proteins are involved in vesicle fusion in every cell of the animal kingdom – a universal mechanism for secretion.
In the course of his experiments, Rothman created a technique for mixing NSF and one of the SNAP proteins together with an extract of brain tissue, and then separating them out again, to see which of several thousands of proteins in the brain extract might have bound to NSF and SNAP. Only three proteins stuck: VAMP/ synaptobrevin, syntaxin and a third called SNAP-25 (which dwells mainly in the plasma membrane). These were all shown previously by Scheller to be involved in neurotransmitter release.
Rothman immediately realized the importance of the result, and proposed the “SNARE hypothesis” for vesicle transport and exocytosis as a general process in all cell types: vesicle proteins specifically bind to target proteins in the plasma membrane to form a "v-t" complex which is necessary for vesicle and membrane fusion to proceed. The "v" and "t" proteins are believed to twist around each other and, in a zipper-like fashion, pull the vesicle and the plasma membrane together until they fuse under the physical force of the twisting proteins to open a pore through which the contents of the vesicle flow to the outside.
“Rothman placed everything in this bigger context. In one stroke he realized that the molecules were involved in membrane trafficking in cell biology in general,” says Charles Stevens of the Salk Institute, a member of the Kavli Neuroscience Committee.
Rothman and Scheller collaborated and proposed that NSF and SNAP form part of the vesicle fusion machinery in neurons, and that synaptotagmin served as a clamp which prevented the fusion process from happening until a calcium signal arrived.
This arrangement means that vesicles can stack up close to the plasma membrane, ready to let out their contents in the milliseconds following the arrival of a nerve impulse, but without emptying too soon.
“You wouldn't have time in the speed required for synaptic transmission to make the vesicles, load them up, and put them in the active zone ready to release them. It’s all there ready to go – the readily releasable pool – like a loaded canon. All you’ve got to do is pull and release,” explains Eric Kandel at Columbia University, and member of the Kavli Neuroscience Prize committee.
Rothman also proposed that SNAP-25, VAMP/synaptobrevin, and syntaxin represent members of a large family of proteins, different forms of which are involved in membrane fusion in all cells, and in different compartments within cells. NSF contributes by providing a shot of energy through splitting the energy-containing molecule ATP, for the scaffold to then break apart ready to reassemble again. Without the energy released from ATP this step would take a billion years according to Rothman (Nature Medicine, vol 8(10), October 2002).
Südhof used genetic engineering to "knock-out" the synaptotagmin gene in mice, with the result that synapses from the forebrains of mice that lacked synaptotagmin lost their capacity for fast calcium-triggered release of neurotransmitter, thus confirming the importance of synaptotagmin in vivo.
Südhof performed X-ray crystallography and other studies to reveal how a different group of proteins, the complexins, bind to the SNARE scaffold and help to stabilise the structure until synaptotagmin displaces them, calcium-activated synaptotagmin then sparking the fusion of the vesicle with the pre-synaptic membrane.
Scheller, Südhof, and other investigators also demonstrated that the bacterial poisons tetanus toxin and botulinum toxin block neurotransmitter release by cleaving apart the vesicle and membrane associated proteins synaptobrevin, SNAP-25, and syntaxin.
“Before the work of Scheller, Südhof, and Rothman, the target for these toxins was known to be at the neuromuscular junction and at the synaptic membrane, but the molecular target was not known,” says Neuroscience Prize Committee member Jean-Pierre Changeux at the Pasteur Institute, Paris.
Scheller went on to use electron microscopy to pin point the location of v- and t-SNARE proteins on vesicles and the pre-synaptic membrane. In 2001 he then moved to the biotech company Genentech, leaving Südhof to continue making major contributions to the field. Südhof also identified two proteins, Rab3 and RIM, which assist in the movement of vesicles towards the pre-synaptic membrane – to the region known as the "active zone."
Recently, Rothman and Südhof described the role of the Rab proteins and the SM (Sec1/Munc18-like) proteins in providing additional specificity and control to the process of SNARE scaffold assembly and vesicle docking to membranes in various cell types, thus confirming the general importance of these proteins (Science, vol 323, 23 January 2009).
Thus Scheller, Südhof, and Rothman have together revealed not only the identity of the molecules that initiate vesicle fusion and neurotransmitter release, but also how they act in a coordinated and precisely controlled manner that ensures neuronal signalling occurs with maximum efficiency and minimum background "noise." Their findings form the basis for future investigation into the finer details of neuronal signalling in the brain and how these relate to processes such as learning and memory, and the defects responsible for conditions such as schizophrenia and autism.
“We knew synaptic transmission was vital and probabilistic, but we knew nothing about the mechanisms. They took a big step towards explaining the molecular mechanism by which it happens and that’s a big deal!” says Committee member Charles Stevens.
“What happens at the membrane in the release of neurotransmitter – this is really fundamental to the nervous system,” says Jon Storm-Mathisen, chair of the Kavli Prize for Neuroscience committee.